STABILITE D’UNE
BANQUE DE DONNEES
SOCIOECOLOGIQUES
Comportements des plantes de la banque
SOPHY en 2001 et en 2008
Liste des taxons indices de variables
La
banque de données phytosociologiques SOPHY a débuté son existence en 1978. Ses
promoteurs n’ont eu de cesse de lui ajouter de nouvelles données et d’en
obtenir de nouveaux résultats. L’un d’entre eux, remarquable par sa nouveauté
est la possibilité de montrer la probabilité
de présence des plantes dans chacun des relevés. C’est en quelque sorte une
façon de montrer les milieux favorables à chacune des plantes de la banque. On
appelle ce type de résultat la « flore
probable ».
Le
premier résultat important concernant la flore probable a été installé sur le
site SOPHY début 2002 à partir des données rassemblées jusqu’en 2001. Il concernait 3700 plantes (5273
en tenant compte des niveaux d’abondance PNA) observées dans 115.000 relevés
localisés. Le résultat actuel (2008)
porte sur 4598 plantes observées (7883 PNA) dans 172.000 relevés soit une
augmentation de la moitié des relevés rassemblés précédemment. C’est pourquoi
il nous a paru intéressant de procéder à la comparaison des comportements
socioécologiques des plantes communes à ces deux stades de la banque de façon à
répondre à la question : l’apport de données nouvelles a-t-il modifié de
façon sensible les résultats obtenus antérieurement ?
La
comparaison ci-dessous porte sur deux aspects de ces résultats : d’une
part, les plantes écologiquement similaires, d’autre part la répartition
géographique de la flore probable. Mais tout d’abord rappelons la façon dont
est déterminée la caractérisation socioécologique des plantes.
1. PRINCIPE
DU CALCUL : Caractérisation
socioécologique d’une plante
1.1.
Fidélités des
plantes aux plantes
Dans
une station où se trouve une plante donnée (A), celle-ci rencontre d’autres
plantes qui vivent ensemble dans le même milieu. Ces plantes sont ses
« co-occurrentes » (C). Les plantes C sont des indices du milieu de
A. Elles servent à l’identifier. Dans l’ensemble des stations de A, plus une
plante C est fréquente, plus elle contribue à caractériser le milieu de A. Ces
principes de base sont adoptés depuis longtemps par toute la phytosociologie. A
cette base, la banque Sophy a naguère apporté une nouveauté, permise par le
développement de l’informatique. Cette nouveauté consiste à appliquer une
notion fondamentale de la phytosociologie, la fidélité, pour mesurer les
relations entre les plantes. Initialement, la fidélité d’une plante à un
groupement permettait d’identifier les plantes caractéristiques du groupement.
Dans une banque telle que Sophy, la fidélité permet aussi de quantifier les
similitudes entre les comportements des plantes. Le comportement écologique
d’une plante se quantifie par ses dépendances apparentes envers les milieux des
autres plantes. La dépendance apparente de la plante A envers le milieu de C se
mesure par la fidélité de A à C, FID(A,C). Cette fidélité est une probabilité.
Elle est égale à la fréquence relative de C dans les relevés contenant A.
1.2. Le tableau carré des
fidélités, source des caractérisations socioécologiques
L’écologie
de la plante A se caractérise par ses fidélités à toutes ses cooccurrentes,
considérées comme des indices du milieu. Elle se caractérise aussi par ses
fidélités nulles aux plantes qu’elle ne rencontre jamais. Au total, la plante A
se caractérise par autant de probabilités qu’il y a d’indices de variable
(7883).
Le
tableau des fidélités des plantes A aux plantes C est un tableau carré puisque
finalement le nombre de cooccurrentes C est égal au nombre de plantes A. Une
ligne de ce tableau contient la caractérisation socioécologique d’une plante A
qui est l’image de son comportement écologique. Une colonne contient les fidélités
des plantes à un indice de variable C. A
partir de ce tableau carré, on peut comparer une plante A à toutes les autres,
d’après leurs comportements. On peut notamment identifier les plantes les plus
similaires de A.
1.3. Limiter l’effet du
suréchantillonnage des relevés
Ce
calcul simple de la similitude a rencontré un écueil, ce qui a nécessité le
perfectionnement du calcul. Cet écueil fut la fidélité exagérée d’une plante à
une autre, du fait de leur coexistence fréquente dans des localités marginales
de leur aire, lorsque ces localités ont fait l’objet d’un suréchantillonnage
détaillé. Pour diminuer l’influence de ce suréchantillonnage, les stations sont
regroupées dans des « quadrats » d’environ 1km de côté, en fait d’un
centigrade, en latitude comme en longitude. Ce regroupement ne confond pas les
relevés d’un même quadrat pour en faire un gros relevé collectif. Le relevé
reste un échantillon de milieu distinct des autres relevés du même quadrat. On
mesure la similitude écologique de deux plantes par leur coexistence plus ou
moins fréquente dans les mêmes relevés, mais on donne la même importance à tous
les quadrats, quelle que soit le nombre de relevés (NR) qu’il contient. Par
conséquent, dans un quadrat, on donne à chaque observation un poids égal à
1/NR. On évite ainsi de donner un poids excessif aux nombreux relevés parfois
répétés côte-à-côte par certaines études botaniques locales.
2. STABILITE
DES PLANTES ECOLOGIQUEMENT SIMILAIRES
2.1. Calcul de la similitude socioécologique entre
deux plantes
Une
plante se caractérise par ses fidélités à des milliers d’indices de
variable : 5273 en 2001, 7883 en 2008.
La différence entre les comportements écologiques de deux plantes se
mesure par une probabilité. Si on
appelle FA et FB les fidélités des plantes A et B à un indice C, la probabilité
de différence entre A et B à l’égard de C vaut│FA-FB│ en valeur
absolue. Cette différence mesure la
disjonction entre A et B par rapport à C. La différence entre les
comportements de A et B d’après tous les indices du milieu est la moyenne des
différences │FA-FB│.
Jadis, un premier calcul utilisait la simple moyenne, et il aboutissait à un
mélange de résultats pertinents et de résultats absurdes. En particulier, les
plantes xérophiles apparaissaient similaires aux plantes hygrophiles ! La
cause de cet artéfact était due aux fidélités également nulles des xérophiles comme des hygrophiles envers
les nombreuses plantes mésophiles. Xérophiles et hygrophiles ont en commun des
caractères négatifs (des fidélités nulles), donc des différences moyennes
faibles. Cet artéfact nous a montré que, dans la différence entre les plantes A
et B, la contribution de C a une pertinence qui dépend de sa rencontre
effective avec A ou avec B. Cette pertinence est égale à la fréquence des
couples de relevés (A, B) qui possèdent C. Cette pertinence vaut (FA + FB – FA
x FB). Si FA et FB sont nulles (comme dans le cas précédent de A xérophile et B
hygrophile), le poids est nul. Si FA ou FB vaut 1, le poids vaut 1. En définitive, la différence entre les
comportements de A et B est la somme de│FA-FB│ divisée par la somme
de (FA + FB – FA x FB), pour tous les indices C.
Ces
différences sont triées par valeurs croissantes. Les différences les plus
faibles désignent les plantes les plus similaires. Les trente premières plantes
suffisent à identifier un cortège floristique typique de la plante (Tableau I).
2.2. Exemple de stabilité
dans les similitudes
Tableau I
Comparaison des plantes socioécologiquement similaires
à Cistus albidus présent (1-6),
à deux stades de la banque SOPHY
|
En 2001 :
présent dans 843 quadrats Nb de cooccurrentes : 1655 Plantes écologiquement similaires PDI A-D COOC .15 ONONIS MINUTISSIMA L. 1-5 2419 1695 .16 PINUS HALEPENSIS MILL. 1-6 5142 1771 .17 CISTUS ALBIDUS L.
3-6 578 1026 .19 JUNIPERUS OXYCEDRUS L. 1-6 5119 2014 .20 PSORALEA BITUMINOSA L. 1-5 2446 2021 .21 THYMUS VULGARIS L. 1-6 2007 2406 .22 AVENA BROMOIDES GOUAN 1-4 4358 2015 .22 JUNIPERUS OXYCEDRUS L. 2-6 5118 1505 .23 SPARTIUM JUNCEUM L. 1-6 2454 1687 .23 PINUS HALEPENSIS MILL. 4-6
5141 1040 .23 DORYCNIUM SUFFRUTICOSUM 1-6 2277 1983 .24 CALYCOTOME SPINOSA (L.) 1-6 2241 1435 .24 RHAMNUS
ALATERNUS L. 1-5 2977 1836 .24 FUMANA
CORIDIFOLIA (VIL 1-4 590 1619 .25
ODONTITES LUTEA (L.) RC 1-5 3509 2162 .25
PISTACIA TEREBINTHUS L. 1-5 2580 1548 .25 ODONTITES LUTEA (L.)
RC 2-5 3508 1717 .25 PSORALEA BITUMINOSA
L. 2-5 2445 1447 .25 SPARTIUM JUNCEUM L. 2-6 2453 1302 .25 LOROGLOSSUM LONGIBRACTE 1-2 5004 986 .25 HELICHRYSUM STAECHAS (L 1-6 954 2282 .26 ROSMARINUS OFFICINALIS 1-6 1925 1707 .28 EUPHORBIA NICAEENSIS .28 SEDUM NICAEENSE ALL. 1-4 1262 2405 .29 ARGYROLOBIUM LINNAEANUM 1-5 2202 1636 .29 PISTACIA
LENTISCUS L. 1-6 2578 1741 .29 CORIS MONSPELIENSIS L. 1-3 1225 1288 .29 BRAC PINN SUBS PHOENICO 1-6 4382 2448 .29 LONICERA IMPLEXA AITON 1-4
239 1752 .29 CLEMATIS
FLAMMULA L. 1-5 2854 2192 |
En 2008 :
présent dans 1063 quadrats Nb de cooccurrentes : 2030 Plantes écologiquement similaires PDI A-D COOC .16 PINUS
HALEPENSIS MILL. 1-6 7478 2221 .18 ONONIS
MINUTISSIMA L. 1-5 3554 2151 .18 CISTUS ALBIDUS
L. 3-6 861 1237 .19 JUNIPERUS OXYCEDRUS L. 1-6 7448 2577 .21 JUNIPERUS OXYCEDRUS L. 2-6 7447 1906 .21
PSORALEA BITUMINOSA L. 1-5 3594 2702 .22 PINUS HALEPENSIS
MILL. 4-6 7477 1267 .23 THYMUS
VULGARIS L. 1-6 2947 3200 .23 AVENA
BROMOIDES GOUAN 1-4 6395 2552 .24 SPARTIUM JUNCEUM L. 1-6 3606 2116 .24 DORYCNIUM SUFFRUTICOSUM 1-6 3350 2587 .25 ONONIS MINUTISSIMA L. 2-5
3553 1640 .25 PSORALEA BITUMINOSA L. 2-5
3593 1957 .25 CALYCOTOME SPINOSA (L.) 1-6 3295 1762 .25 RHAMNUS ALATERNUS L. 1-5 4399 2422 .26 FUMANA CORIDIFOLIA (VIL 1-4 882 2143 .26 ROSMARINUS OFFICINALIS 1-6 2838 2192 .26 PISTACIA TEREBINTHUS L. 1-6 3786 1905 .26 ODONTITES LUTEA (L.) RC 1-5 5128 2856 .26 HELICHRYSUM STAECHAS (L 1-6 1421 2984 .27 ODONTITES LUTEA (L.) RC 2-5 5127 2244 .27 SPARTIUM JUNCEUM L. 2-6 3605 1584 .28 LOROGLOSSUM LONGIBRACTE 1-2 7292 1134 .29 FUMANA THYMIFOLIA (L.) 1-5
888 1400 .29 CALYCOTOME SPINOSA (L.) 3-6 3294
999 .30 CORIS MONSPELIENSIS L. 1-3 1813 1574 .30 ARGYROLOBIUM LINNAEANUM 1-5 3229 2086 .30 SEDUM NICAEENSE ALL. 1-4 1866 3141 .30 BRAC PINN SUBS PHOENICO 1-6 6427 3115 .30 EUPHORBIA NICAEENSIS AL 1-4 2396 1437 |
Légende :
en bleu et en noir, les plantes communes aux
deux listes ; en vert, les plantes qui
diffèrent
En bleu
et en vert, les plantes munies d’un lien
HTML ; en noir, celles qui en
sont dépourvues.
PDI est la
probabilité de différence entre les deux plantes ; COOC, le nombre de cooccurrentes ;
A-D les
limites des classes d’abondance-dominance : ex. Pinus halepensis est représenté au niveau de la simple présence
(abondances comprises entre 1 et 6), et
par un autre niveau aux abondances comprises entre 4 et 6.
Dans
cet exemple, la plupart des plantes écologiquement similaires sont communes aux
deux stades de la banque : en outre, les valeurs des probabilités de différence entre deux plantes restent très
voisines. Il y a simplement quelques décalages dans le classement. Cette grande
stabilité est due à l’importance du nombre des relevés effectués dans toute
3. STABILITE
DE
3.1. Probabilité de présence d’une plante dans un
relevé
Une plante C, présente dans un relevé, indique une
probabilité d’y trouver aussi la plante A. Cette probabilité, on l’a vu, est la
dépendance apparente de C envers le milieu de A, autrement dit la fidélité de C
à A. Comme chaque plante C apporte sa propre information, l’information
résultante est donnée par la moyenne des fidélités des plantes du relevé envers
A pour toutes les plantes du relevé. Toutefois, le calcul de la fidélité
moyenne envers A ne tient pas compte de la présence éventuelle de A dans le
relevé. En effet, cette présence apporte une indication qui n’est plus une
probabilité mais plutôt une tautologie, puisque la fidélité de A à A vaut 1.
Cette valeur est très supérieure à la plupart des fidélités des autres plantes
envers A et elle éclipserait leur message. La fidélité moyenne des relevés
envers A permet de cartographier la présence probable de A dans tous les
relevés, y compris ceux dont A est absente.
3.2. Cartographie de la
flore probable
Pour
mettre en évidence les gradients de la flore probable, il faut préalablement
trier les valeurs des probabilités de présence d’une plante (des plus fortes
aux plus faibles) puis les répartir en classes. Six classes d’effectif cumulé
croissant sont représentées : 2%, 5%, 10%, 20%, 50% et 100% de l’effectif
total des valeurs non nulles. Ces effectifs suivent une progression géométrique
simplifiée. Aux probabilités les plus
fortes sont attribuées les couleurs les plus foncées, aux probabilités les plus
faibles les couleurs les plus claires. Le noyau des probabilités maximales est
souvent géographiquement concentré. Il est englobé par les classes suivantes
comme dans un halo. Pour rendre apparente la structure géographique de cette
gradation, on cartographie un nombre de localités qui dépend du nombre de
relevés possédant la plante. Pour plante peu fréquente, on représente un nombre
de localités supérieur au nombre de relevés ; pour une plante fréquente on
représente un nombre de localités inférieur au nombre de relevés où elle est
observée. Par exemple, Aristolochia
altissima est observé dans 13 quadrats mais représenté par 252 quadrats, Cistus albidus est observé dans 1063
quadrats et représenté par 2282 quadrats, Sorbus
aucuparia, 4843 et 4871, Hedera helix,
11195 et 7406.
3.3. Exemples de
répartitions probables
|
|
|
|
en 2001 |
en 2008 |

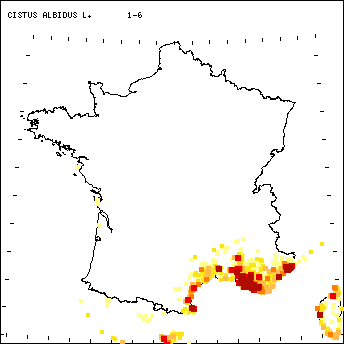
Figure 1.- Répartition des
milieux favorables à Cistus albidus
aux deux stades de la banque SOPHY
Commentaires de la carte.
Le
territoire dans lequel s’inscrit la probabilité de présence de Cistus albidus est relativement continu.
Un gradient écologique est perceptible principalement du Sud vers le Nord. A
noter que Cistus albidus est probable
en Corse alors que sa présence (non montrée ici) est très réduite. Il est aussi
très faiblement probable sur le littoral atlantique alors qu’aucun relevé de ce
secteur ne le contient. Pour une plante de cette fréquence, la localisation des
milieux favorables à Cistus albidus
n’a guère changé entre ces deux stades.
4. TROIS CONCLUSIONS : EXPRESSION PROBABILISTE,
COHESION SOCIOECOLOGIQUE,
STABILITE DES RESULTATS
4.1. Expression probabiliste
La probabilité est la forme générale de la
connaissance sur le comportement des plantes en milieu naturel. Dans ce
travail, les critères quantitatifs sont définis directement comme des
probabilités, que ce soit la dépendance apparente d’une plante envers une
autre, ou bien la similitude entre deux comportements, ou encore la probabilité
de présence d’une plante dans un relevé. La probabilité est un calcul simple
comportant peu d’artéfacts et peu de déformation systématique, contrairement à
l’arsenal de la statistique géométrique (corrélations, régressions, analyses
factorielles).
4.2. Cohésion socioécologique
Le tableau des fidélités mutuelles des plantes aux
plantes permet de relier tous les comportements les uns par rapport aux autres
et de comparer notamment les comportements des espèces qui s’excluent
mutuellement, ou encore les comportements des espèces voisines, dont les études
locales ont du mal à peser les nuances, alors que la socioécologie les
différencie clairement. Le tableau des fidélités permet aussi de situer les
relevés les uns par rapport aux autres, de les hiérarchiser en groupements, de
les caractériser par leurs plantes discriminantes. Ce tableau est le cerveau de
la banque.
4.3. Stabilité des résultats
L’effectif nombreux et la précision systématique des
données stockées dans la banque SOPHY apportent deux sortes de stabilité. En
premier lieu, on vient de le montrer, les caractérisations de la plupart des
plantes restent stables alors que la banque s’enrichit. Cet enrichissement n’en
reste pas moins nécessaire pour stabiliser les comportements des plantes les
moins fréquentes ou les moins bien échantillonnées. En second lieu, les
synthèses peuvent être répétées, presque à l’identique, à partir de lots distincts
de données, comme nous l’avons montré dans un autre travail qui portait sur les
groupements végétaux de